What do an elephant trunk, a cephalopods tentacles, and the mammalian tongue have in common? They are all examples of muscular hydrostats.
In a muscular hydrostat the muscle fiber orientation and lack of internal bony structure, or bony origins and insertions, allow freedom of movement and deformation making them great for manipulating items, including food. The free movement of the tongue facilitates its use during feeding and drinking, and in humans, for speech. One expectation of a muscular hydrostat is that for changes in one dimension (i.e. shortening) there is simultaneous compensation on other dimensions (i.e. widening and/or thickening). While most muscles can be readily modeled, the tongue’s freedom of movement and deformation make modeling and interpreting its dynamic functional properties difficult.
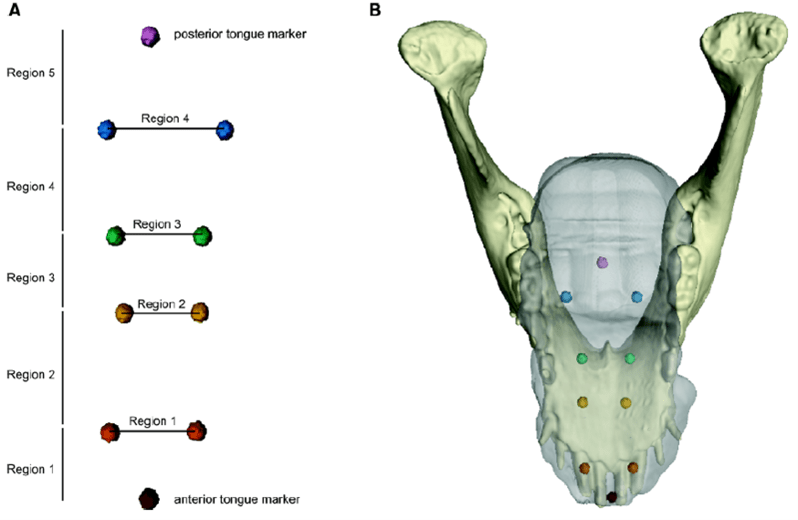
Olson et al (2021) are taking a deeper dive into the biomechanics of the tongue to explore the finer scale movements during drinking and feeding. To do this, they used a combination of techniques including computed tomography (CT) and X-ray Reconstruction of Moving Morphology (XROMM). CT data lets researchers model 3-D shapes of structures (the 3D mandible model in Figure 1) and the XROMM data let researchers follow the internal movement of bony structures during different actions. For XROMM, they used markers in the jaw, skull, and the tongue (Figure 1), these hard and soft tissue markers enabled comparison of tongue and jaw movements.
In Figure 2 we can trace jaw pitch (angle of jaw opening) along with the change in length of the tongue in region 3 (green points in Figure 1). In Figure 2 the jaw is closed at the central peak of the figure and opened at the beginning and end of the trace. Open and close your jaw to see what your tongue does as you go through a cycle!

Following expectations of the muscular hydrostat, the maximum tongue length coincided with narrowing while minimum length coincided with tongue widening. However, there were also differences between chewing and drinking and among the tongue regions. During chewing, the tongue is deforming more than in drinking, but overall, the tongue is wider when the pigs are drinking. By region, the posterior part of the tongue is deforming more than the anterior part while also deforming later in the gape cycle.
With differences in the deformation timing across regions, the pigs maintain similar total tongue length throughout the gape cycle (a single opening/closing of the jaws). If all regions of the tongue deformed together, the researchers would have seen greater changes in total tongue length. Chewing and drinking also differ in the timing of tongue deformation compared to timing of the gape cycle, not something the research team expected. Differences between chewing and drinking may highlight functional differences necessary for these behaviors. When drinking, the tongue is facilitating fluid transport while in chewing, it facilitates food placement for mastication.
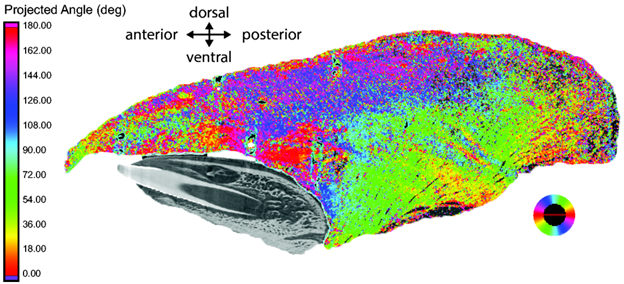
So far we’ve seen the differences in fine-scale movements and deformation of the tongue, but how are the pigs able to do this? To understand how the tongue’s morphology supports its function, Olson and colleagues used CT scanning with iodine staining to visualize the muscle fiber orientation (Figure 3). The complex muscle fiber arrangement supports the regional differences found during feeding and drinking in pigs.
This study presents critical exploration of regional differences in tongue deformation during chewing and drinking. It has laid the groundwork for work on other aspects of how the tongue functions during different behaviors.
Dr. Callie Crawford is a postdoctoral fellow at the University of Louisiana – Lafayette. Read more about her here – https://crawfordcallie.wixsite.com/home.
