We paleontologists insist that our field is more than a depressing inventory of prehistoric loss motivated by a morbid taste for bones. Instead, we argue that paleontology is key to a complete understanding of the natural world. To live up to this argument, paleontology must accurately reconstruct how past organisms lived; once this is accomplished, the many past Earths of prehistory can be explored. In this paleontological view of life, the present world and its inhabitants are a small minority: most things that have ever lived have gone extinct, and most worlds have been lost. At its best, paleontological reconstruction and exploration of the past enables biology to reveal the rules of life on Earth, not just the rules of life on the present Earth.
The paleontological reconstruction of past worlds is a big topic best broken down into smaller chunks. A key component is comparative biomechanics, the art of inferring how a target organism worked and performed through comparison to other organisms. In the paleontological reconstruction of extinct organisms, comparative biomechanics varies in effectiveness depending on the sort of extinction involved. If extinction were rare, minor, and evenly distributed, very little information would be lost and paleontological reconstruction would be easy. In the classic metaphor where biological diversity is represented by a vast tree with major groups for boughs and species for twigs, this would be like infrequently removing a single twig from each bough – the overall shape of the tree would not be affected. In this best case for comparative biomechanics, the species represented by each twig could easily be reconstructed by comparison with neighboring twigs. But this is not usually how extinction occurs. Instead, extinction is common, significant, and unevenly distributed; and sometimes even the most successful and resilient groups are eliminated all at once in mass extinctions. This sort of extinction alters the path of history: the loss of entire boughs irreversibly changes the shape of the tree. In this case, closely related species (or neighboring twigs on the tree of life) are necessarily lost too. These losses can be a big problem for comparative biomechanics depending on the shape of the tree of life after it is trimmed.
Sometimes, the surviving groups flanking the extinct group on the tree of life are similar enough (both in evolutionary relatedness and anatomy) to allow meaningful comparison. This sort of analysis would be like a future archaeologist attempting to reconstruct the function of a broken electric mixer through comparison to an intact whisk and a blender. This approach is exemplified by paleontological reconstruction of dinosaurs using comparisons to alligators and birds. Alligators split from the broader group that contains dinosaurs before dinosaurs themselves split off as their own group. Later, birds split off from a specific group within dinosaurs (Figure 1a). If a structure functions in the same way in both alligators and birds, it is reasonable to expect that the structure would have functioned in a similar way in dinosaurs.
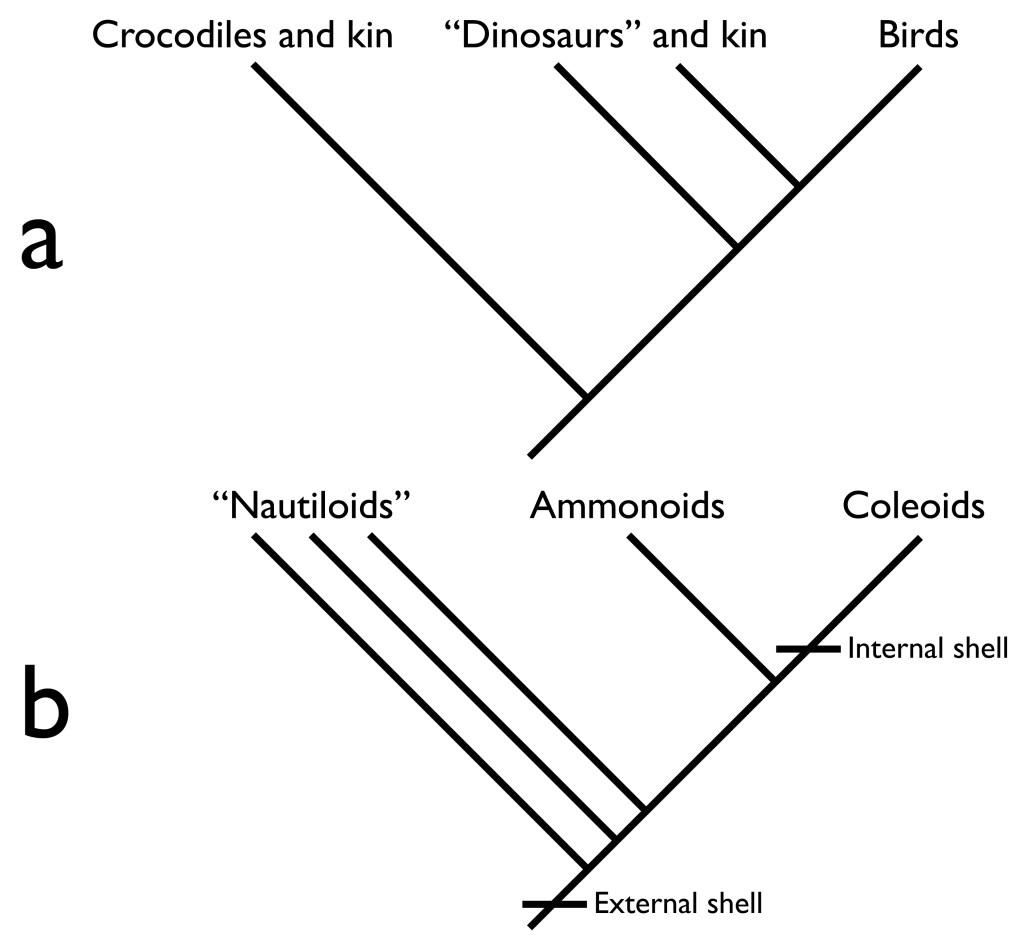
But sometimes the living relatives of a group are simply too different for useful comparisons, and comparative biomechanics becomes ineffective. For example, ammonoids (Figure 2a) were a group of coiled cephalopod mollusks related to coleoids, another group of cephalopods including squids, octopus, and cuttlefish (Figure 1b). Ammonoid fossils indicate they would have relied on the same water-jet propulsion system as coleoids, but that is where the functional similarity ends. This is because ammonoids have external shells (making them ectocochleates, the umbrella term for externally shelled cephalopods), unlike their naked coleoid kin. This difference alone means that the details of ammonoid behaviors and functions (e.g., swimming performance) cannot be reconstructed from coleoid performance. This is especially troublesome because ammonoids were an incredibly successful group and key components of ancient ecosystems. They lasted more than 340 million years from their first appearance in the Devonian period before being wiped out (along with the dinosaurs) during a mass extinction at the end of the Cretaceous period. It’s worth pointing out that their extinction had nothing to do with obsolescence or inadequacy inherent to external shells – ammonoids survived mass extinctions and recovered to flourishing multiple times over their long history. They simply ran out of luck at the end of the Cretaceous.
There is one surviving group of externally shelled cephalopods: the nautiloids (Figure 2b). Unfortunately, these are not particularly closely related to ammonoids – they are the last surviving members of a diverse set of animals that split off from the ancestor of both coleoids and ammonoids. Furthermore, there are only a few types of living nautiloid with minor differences in shell anatomy, unlike the diverse ammonoids and their wide variation in shell construction and coiling. A comparative biomechanical approach relying on nautiloids can only go so far. Nautiloids can provide a guide to how ectocochleates work in general, but cannot provide critical information for reconstruction of ammonoids, such as how differences in ammonoid shell anatomy would affect their performance and influence how they lived. This creates a tricky situation for paleontologists. The surviving groups close to ammonoids on the tree of life are very different anatomically; the only surviving group similar to ammonoids in anatomy is very distant on the tree of life. Forget the simple comparison between whisks, blenders, and electric mixers. This scenario would be like a future archeologist attempting to reconstruct the performance of winged aircraft across the history of aviation – from the Wright Flyer to the Avro Arrow – from a single model of craft produced by a single manufacturer. So, although the reconstruction of ammonoid function and behavior may seem like a niche issue at first, it strikes at the heart of the entire field of paleontology. If paleontology is to enable biology to understand life on Earth and its entire range of possibilities (not just those represented in the present), it must be able to overcome extinction. It must be able to reconstruct the past lives of extinct organisms, including animals very different from those living today (such as ammonoids) and other cases where comparative biomechanics is ineffective.

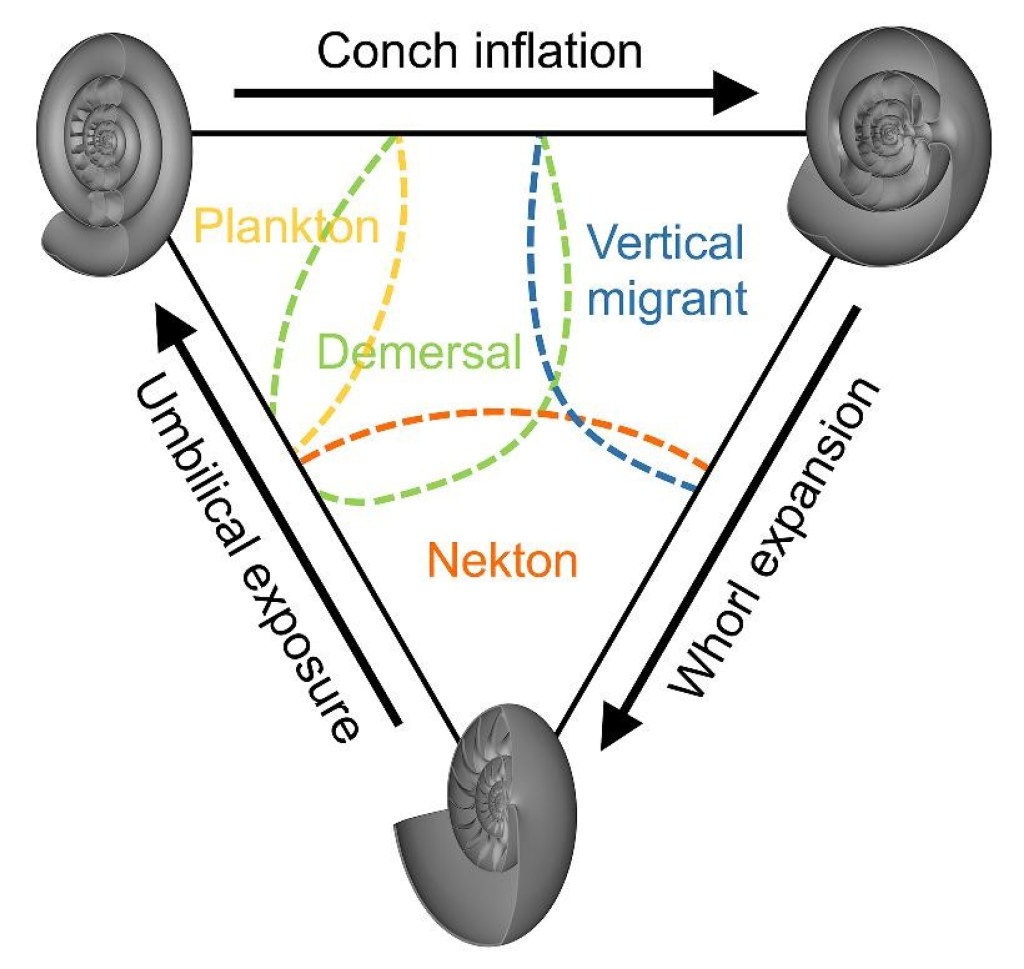
In an attempt to meet this challenge, previous approaches to ammonoid biomechanics came up with reasonable hypotheses based on broad anatomical categories (Figure 3). The range of variation in ammonoid shells can be expressed in a triangular diagram, with three categories – oxycone, serpenticone, and spherocone – as the extreme anatomies forming its corners. Oxycone shells are narrow and streamlined side-to-side. Their outermost rings are larger than the central rings, making it so that most of the shell is made up by the outer ring, which also aids in streamlining. This combination of traits leads to the hypothesis that these animals were active swimmers capable of moving against the current. The rings of serpenticone shells are closer to circular in cross-section and have only a gradual increase in size from the inside to outside rings. This configuration is hypothesized to result in lower swimming performance and leads to interpretation of these animals as drifting with the current. Finally, spherocone shells have extremely wide bodies, while sharing swollen outer ring anatomy with oxycone shells. These animals are interpreted as primarily concerned with vertical movement up and down in the depths. Some early experiments demonstrated that oxycone and serpenticone shells had lower drag and were better at coasting but could not consider other critical aspects of swimming performance, such as maneuverability, leaving the biomechanical hypotheses and biological interpretations mostly untested. Without good living comparisons for ammonoids, a new approach to hypothesis testing was needed.
Two scientists at the University of Utah – David J. Peterman and Kathleen A. Ritterbush – rose to the challenge of testing these hypotheses more fully. Instead of relying on comparisons with living species, Peterman and Ritterbush used advanced, realistic models of oxycone, serpenticone, and spherocone shells to test how they would perform in life. Nautiloids still had a role: to ground truth the experiment, a model nautiloid shell was constructed using the same method as used for the ammonoid shells. In a compelling example of how science progresses, this was the latest in a series of experiments – each building on the last – that previously included other scientists, such as Christopher C. Barton, David J. Bottjer, and Margaret M. Yacobucci. These past experiments generated 3D virtual models of ammonoids, calculated important metrics related to swimming performance, and produced 3D-printed robots. A first test involving the 3D-printed robots revealed that spherocone shells had superior performance in side-to-side maneuverability (yaw maneuverability) but low side-to-side stability (hydrodynamic stability) but could not determine the up-and-down maneuverability (pitch maneuverability) or up-and-down stability (hydrostatic stability). The latest experiment (our focus here) adds a major innovation allowing pitch maneuverability and hydrostatic stability to be examined: counterweights were carefully calibrated and positioned within the model shells. This adjusted the center of mass (the point around which the mass of the animal is evenly distributed) relative to the center of buoyancy (the center of mass for the body of water displaced by the animal). This is a critically important parameter. If an ammonoid is to be stable in the water – and not to spin like an out-of-control wheel – its center of mass must be below its center of buoyancy. The lower the center of mass relative to the center of buoyancy, the more stable the animal. Different shell shapes influence how the soft body of the animals sits in the shell and therefore its center of mass (Figure 4), so the counterweights had to be positioned differently, based on previous calculations, for each of the oxycone, serpenticone, and spherocone shells.

Once the counterweights were positioned correctly, Peterman and Ritterbush used high-resolution cameras and 3D-motion tracking technology to record the pitch maneuverability and hydrostatic stability of the model ammonoid shells. The model ammonoid shells were rotated away from their stable, upright position (equilibrium) then released remotely using a robotic arm. The model ammonoid shells oscillated back and forth like a pendulum but varied in the speed at which they rotated and how quickly they restored themselves to equilibrium. More stable model ammonoid shells were expected to rotate faster – undergoing more oscillations in the same amount of time – and restore themselves to equilibrium more quickly. Plotted on a graph, more stable ammonites were expected to have more, smaller oscillations than less stable ammonites in the same amount of time. Like most things in life, however, this came with a trade-off. More stable ammonite shells would necessarily be less maneuverable, as their stable shells would also resist deliberate attempts by the animal to rotate away from its equilibrium position.
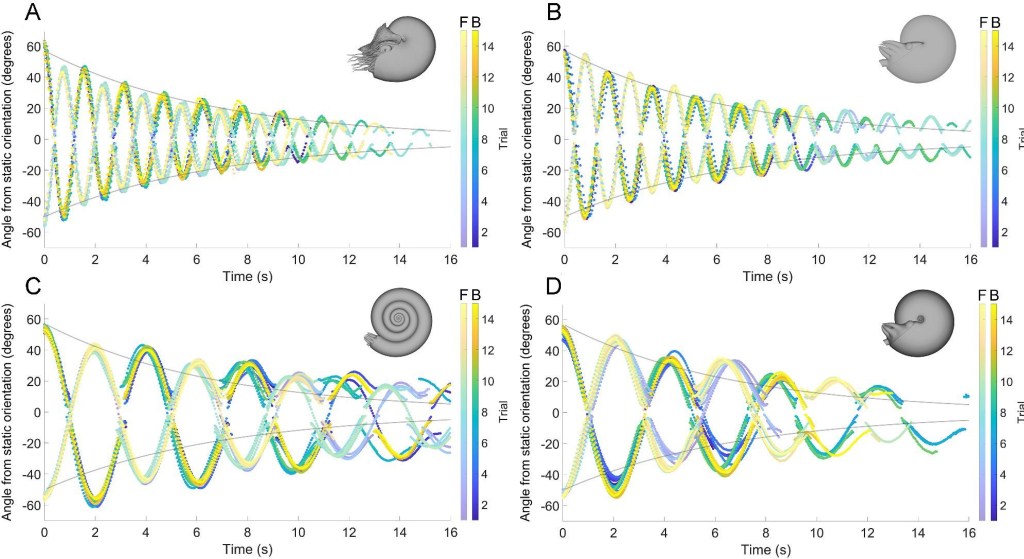
A clear pattern emerged once all the shells were tested and the data plotted (Figure 5). The spherocone and serpenticone shells, with their evenly distributed whorls, took longer to slow their oscillations and restore themselves to equilibrium. This implied low hydrostatic stability but high pitch maneuverability. The oxycone shell, with its large outer whorls, slowed its oscillations and restored itself to equilibrium more quickly. This implied high hydrostatic stability but low pitch maneuverability. But the highest hydrostatic stability and lowest pitch maneuverability were found in the model nautiloid, reinforcing the idea that nautiloids are unique (thanks to millions of years of independent evolution) and not representative of most ammonoids.
Nevertheless, the performance data from nautiloids can be combined with data from ammonoids to develop the big picture of how coiled external shells affect how organisms swim. A combination of traits ensures the hydrostatic stability of nautiloids: relatively large outer whorls and wide bodies. The large outer whorls lower the center of mass relative to the center of buoyancy, ensuring the nautiloid model rotates preferentially towards its equilibrium position. As the animal rotates, the wide body produces drag, parachuting the animal to a stop. Oxycone ammonoids have low centers of mass but narrow bodies, spherocone ammonoids have wide bodies but relatively high centers of mass, and serpenticone ammonoids couple relatively high centers of mass with narrow bodies; none replicate the high stability of nautiloids.
This new understanding of how coiled shells work can be combined with what is known from previous experiments, allowing the triangular diagram expressing ammonoid shell anatomy to be updated with a series of functional tradeoffs (Figure 6). Other than nautiloids, oxycone shells represent the most stable configuration, both in yaw and pitch. This makes sense for an active swimming animal using jet propulsion to swim against the current – powerful thrust without high stability would send the ammonoid into a wild spin – and matches up with the streamlined configuration of oxycone shells. Indeed, oxycone shells seem to be the best suited to controlled travel in every direction (including vertical migration). By contrast, spherocone shells represent the most maneuverable configuration in yaw and pitch. This makes the most sense for an animal engaged in short movements in every direction – spinning to capture food around it in all directions or making quick directional changes to evade predators. However, the lack of stability does not match up with expectations for an animal engaged in vertical migration, as previously hypothesized, since most directions of thrust would throw the animal into a spin. Finally, serpenticone shells marry pitch maneuverability with hydrodynamic stability. Stable movement in a desired direction could probably be achieved if the animal carefully controlled the direction of thrust to minimize vertical rocking.

Of course, most ammonoids have shell anatomies intermediate between the extreme oxycone, serpenticone, and spherocone configurations modelled in this experiment. But determining the pitch/maneuverability tradeoffs experienced by these extreme configurations means that the limits of anatomy can be matched up with the limits of performance; intermediate anatomies can be inferred to have intermediate performance. By experimentally determining limits and functional tradeoffs this mode of reconstruction takes us from speculation of what an animal did to a testable model of what an animal could do. It defines the realm of the functionally possible, which can then be compared with other evidence in the fossil record to narrow things down even further. For example, oxycone shells are often found in rocks produced by fast-flowing water, an environment which seems to match their stable and stream-lined anatomy. This approach can be refined in future work – experiments incorporating intermediate shell anatomies and building models with the elaborate ornamentation and surface features borne by many ammonoid shells could be revealing. Still, this experiment represents a major step forward: it overcomes extinction and the limitations of the comparative method in ammonoid reconstruction by using realistic 3D models. Here, paleontology achieves its ambition and provides an essential contribution by reconstructing and exploring the possibilities of life on Earth.
By Conrad D. Wilson
Conrad D. Wilson is a PhD student in the Department of Earth Sciences at Carleton University, Ottawa. His research focuses on the evolution of functional innovations in early ray-finned fishes. He can be contacted on Twitter (@conradmacwilson) or at conraddwilson@gmail.com.
